РАСПИСАНИЕ ЗАНЯТИЙ НА НЕДЕЛЮ: 16.01.23Г.-20.01.23
| Пт.20.01 | ||||||
1,2 | 306 | 508 | 406 | 505 | 401 | ||
3,4 | 401 | 505 | 505 | 501 | 505 | ||
5 | 401 | 505 | 401 | 306 | - | ||
6,7 | 408 | 501 | - | 508 | 501 |
Здравствуйте, уважаемые студенты, записывайте дату, тему и выполняйте необходимые записи(ВСЁ подряд не пишите, читайте, выбирайте, можно составить план, ЕСЛИ ЕСТЬ ВИДЕО, НАДО ПОСМОТРЕТЬ ,ВЫПОЛНИТЬ ПО НЕМУ ЗАПИСИ, МНОГО НЕ НУЖНО ПИСАТЬ. Материала может быть выложено много, но это не значит, что всё надо записывать! После этого, сфотографируйте и отошлите мне на почту rimma.lu@gmail.com . Тетрадь привезете, когда перейдем на очную форму обучения.)Справа находится АХИВ БЛОГА , смотрите дату и номер своей группы
Моя почта : rimma.lu@gmail.com Жду ваши фотоотчеты!
ГРУППА 306 БИОЛОГИЯ 46, 47
ТЕМА 46,47: Естественный отбор -направляющий фактор эволюции. Формы естественного отбора в популяциях.
Случайные изменения частот генов и генотипов в популяции.
(ПРОЧИТАЙТЕ, СОСТАВЬТЕ КРАТКИЙ ПЛАН )
Естественный отбор выступает в роли главного фактора преобразования организмов,
способствуя выживанию и оставлению потомства наиболее приспособленными особями.
Однако формы естественного отбора, в зависимости от его направленности, эффективности и
особенностей среды обитания организмов, могут быть разными.
Популяции (или виды) состоят из разнообразных организмов, но имеют некоторое среднее
значение любого признака. Это можно представить графически, если по оси Х отложить
значения признака от меньшего к большему, а по оси У - частоту носителей этого признака:

Графическое отображение естественного отбора
График – кривая распределения - имеет форму параболы и отражает нормальное
распределение признака, т.к. очевидно, что большинство особей в популяции имеют среднее
значение признака, а особи с крайними значениями представляют собой меньшинство.
В зависимости от того, где на кривой располагаются наиболее приспособленные фенотипы,
учёные выделяют несколько форм естественного отбора.
Движущий отбор – отбор, при котором сдвиг в ряду поколений происходит в определённом
направлении. Одностороннее давление движущей силы естественного отбора (с одной
стороны) направлено на крайние формы фенотипов.

Движущий отбор
Классическим примером, доказывающим существование движущей формы отбора,
является индустриальный меланизм, описанный на примере изменения
покровительственной окраски в популяции берёзовой пяденицы в Англии в XIX в.
Вследствие развития промышленности стволы деревьев значительно потемнели, а также
погибли светлые лишайники, обитающие на них, из-за чего светлые бабочки стали лучше
видны для птиц, а тёмные — хуже.

Индустриальный меланизм у бабочек
Итак, движущий отбор осуществляется при изменении условий окружающей среды или
приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные
изменения в определённом направлении, перемещая соответственно и норму реакции.
Отбор всегда осуществляется по фенотипам, но вместе с ними отбираются генотипы,
которых их обусловливают.
Любое приспособление всегда относительно. Приспособленность организмов к новым
условиям не означает, что отбор прекращает своё действие в популяции. В постоянных
условиях среды действует стабилизирующий отбор. Эта форма отбора направлена в пользу
среднего значения признака, установившегося в популяции, то есть особи, существенно
отклоняющиеся от среднего значения признаков, устраняются.

Стабилизирующий отбор
Например, во время бури гибнут птицы с короткими и длинными крыльями, а со средними
чаще выживают.
Сходство всех особей, наблюдаемое в популяциях – результат действия стабилизирующей
формы естественного отбора.
Таким образом, стабилизирующая форма отбора в течение сотен тысяч и миллионов
поколений оберегает виды от существенных изменений, от разрушающего влияния
мутационного процесса, выбраковывая мутантные формы. Без стабилизирующего отбора не
было бы устойчивости (стабильности) в живой природе. Стабилизирующий и движущий
отборы взаимосвязаны и представляют две стороны одного процесса. Популяции постоянно
вынуждены приспосабливаться к изменениям условий среды. Движущий отбор будет
сохранять генотипы, которые наиболее соответствуют изменениям среды. Когда условия
среды стабилизируются, отбор приведет к созданию хорошо приспособленной к ней формы.
С этого момента в действие вступает стабилизирующий отбор, который будет поддерживать
типичные, преобладающие генотипы и устранять от размножения уклоняющиеся от средней
нормы мутантные формы.
Для многих популяций характерно существование двух или нескольких форм по тому или
иному признаку – полиморфизм. Его нельзя объяснить только возникновением новых
мутаций. В одних случаях он может быть обусловлен повышенной относительной
жизнеспособностью гетерозигот. В других − может быть результатом действия особой
формы отбора, получившей название дизруптивного, или разрывающего

Дизруптивный отбор
Этот отбор действует в тех случаях, когда две или более генетически различные формы
обладают преимуществом в разных условиях, например в разные сезоны года. Хорошо
изучен случай с преимущественным выживанием в зимний сезон красных, а в летний −
чёрных форм двухточечной божьей коровки. Ещё одним примером действия дизруптивного
отбора может служить существование сезонных рас у ряда сорных растений (например, у
погремка лугового). Дизруптивный отбор благоприятствует более чем одному фенотипу и
направлен против средних промежуточных форм. Он как бы разрывает популяцию по данному признаку на несколько групп, встречающихся на одной территории, и может при участии изоляции привести к разделению популяции на две и
более, т.е. к видообразованию.
Известно, что у самцов многих видов ярко выражены вторичные половые признаки – яркие
перья попугаев, хвост павлина, алые гребни петухов, окраска тропических рыб, бивни
индийского слона и др. Это явление называют половым диморфизмом. На первый взгляд
эти признаки носят неадаптивный характер и осложняют жизнь, делая особи более
заметными для хищников, уменьшая их шансы на выживание. Но выживание – не
единственный компонент приспособленности организмов, второй компонент – успех в
размножении. Ещё Ч. Дарвин обратил внимание на то, что эти компоненты часто вступают
в противоречие. Он же предложил называть отбор, направленный на успех в размножении
половым отбором.

Вторичные половые признаки у самцов разных видов
Самец, который живёт недолго, но нравится самкам, имеет больше шансов на успешное
размножение, так как производит больше потомков, чем тот, который живёт долго, но
оставляет мало потомков. Например, успех самца-павлина у самок напрямую зависит от
яркости его оперения и числа глазков на хвосте.
Рассмотрев формы естественного отбора, можно сделать вывод о том, что он проявляет
творческую роль, отбирая и накапливая полезные для популяции и вида наследственные
изменения и отбрасывая вредные. Это позволяет постепенно создавать новые, более
приспособленные к среде обитания виды.
ТЕМ: Случайные изменения частот генов и генотипов в популяции (Генетическое равновесие в популяции и его нарушение. Изоляция, миграция, дрейф генов, волны жизни).
— Под действием естественного отбора эволюционные изменения популяций всегда сопровождаются направленными изменениями генофонда. Вследствие естественного отбора в популяциях закрепляются полезные гены, то есть благоприятствующие выживанию в данных условиях среды. Доля таких генов возрастает, и общий состав генофонда меняется.
— Причиной изменений генофонда популяций может быть не только отбор, но и другие факторы.
— Рассмотрим их, определим их значение в процессе эволюции.
Для выявления закономерностей, которым подчиняются изменения генофонда, нужно знать, что происходит с частотами аллелей и генотипов в различных условиях. Для начала необходимо ответить на вопрос, как будут изменяться эти частоты в идеальных условиях: численность популяции бесконечно большая, скрещивание всех особей внутри популяций равновероятно и не испытывает никаких затруднений, популяция изолирована, в популяции у особей не возникают новые мутации, естественный отбор не действует.
Сохраняется ли генофонд, таких популяций постоянным или он изменяется?
В 1908 году английский математик Г.Г. Харди немецкий врач В.Р. Вайнберг независимо друг от друга сформулировали и математически доказали, что в популяциях, живущих изолировано, в условиях слабого давления естественного отбора устанавливается генетическое равновесие, то есть наблюдается постоянство частот аллелей различных генов.
Частоты генов, а соответственно и генотипов остаются неизменными из поколения в поколение — это одно из основных положений закона Харди-Вайнберга (напомню, действует только в идеальных популяциях).
Ясно, что ни одна реальная популяция не удовлетворяет этим условиям. Все популяции имеют конечную численность. Обычно они не полностью изолированы друг от друга, каждая популяция неоднородна внутри себя, все гены мутируют и многие мутации имеют селективное значение.
Процессы, обусловливающие “неподчинение” популяции закону Харди-Вайнберга и нарушающие ее генотипическое равновесие, становятся факторами эволюции. Один из этих факторов — мутационный процесс — мы уже рассмотрели.
К числу других факторов, имеющих важное эволюционное значение, относятся изоляция популяций, колебания их численности (“волны жизни”), генетико-автоматические процессы (дрейф-генов).
Необходимую предпосылку для действия всех этих факторов создает скрытое генотипическое разнообразие популяций. Скрытое генотипическое разнообразие в популяциях имеет большой эволюционный смысл: оно позволяет запастись такими вариантами изменчивости, которые обеспечивают быструю адаптацию организмов при появлении в среде новых факторов и при резких изменениях экологической обстановки.
Рассмотрим это на конкретном примере.
Показательный пример возможного использования скрытого генотипического разнообразия (мобилизационного резерва изменчивости) дают популяции насекомых, вырабатывающие устойчивость к инсектицидам.
В нашей стране ДДТ (дихлордифенилтрихлорэтан) впервые был применен в 1942 г. в г. Москве для борьбы с комнатной мухой, а уже в 1946 г. появились первые сообщения о возникновении устойчивых линий этой мухи. Потом оказалось, что регулярное и широкомасштабное использование любого инсектицида в течение нескольких лет обязательно приводит к появлению линий, отличающихся высокой устойчивостью к этому препарату. Конечно, химический яд — это сильнейший фактор отбора. Но учитывая низкую частоту мутаций и их случайный неприспособительный характер, кажется маловероятным, чтобы каждый раз в нужном месте и всего лишь за несколько лет появлялись удачные мутации, обеспечивающие устойчивость к этому фактору. Более вероятно, что естественный отбор извлекает из богатого мобилизационного резерва такие аллели, которые могут быть использованы для создания устойчивости насекомых к ядохимикату.
Итак, изоляция “волны жизни”, дрейф генов, миграция — причины нарушения генетического равновесия, факторы, изменяющие частоты аллелей и генотипов в популяции.
Генетико-автоматические процессы (дрейф генов)
Дрейф генов - случайное ненаправленное изменение частоты генов в популяции.
Дрейф генов наблюдается в малых популяциях, где вероятность случайности велика. Эти различного рода случайности и определяют дальнейшую судьбу генофонда малой популяции.
Это было доказано экспериментально. В пробирки с кормом посадили по две самки и два самца мух дрозофил, гетерозиготных по аллелю А (Ад). В такой искусственно созданной популяции соотношение нормального и мутационного аллелей было одинаковым. Спустя несколько поколений оказалось, что частота мутационного аллеля меняется случайным образом. В некоторых популяциях он был утрачен, в других, наоборот, все особи стали гомозиготными по мутационному аллелю, часть популяции содержала нормальный и мутационный аллели.
Дрейф генов малой популяции может привести к ее исчезновению, но может и способствовать большей приспособляемости ее к среде.
“Волны жизни” и их роль в эволюции
“Волны жизни” (популяционные волны)— периодическое чередование подъемов и спадов численности популяций (термин введен в 1905 г.
С.С. Четвериковым).
Причины колебания численности популяций:
а) хищничество;
б) вспышки эпидемий;
в) засухи, пожары, наводнения и другие природные катастрофы;
г) освоение организмами новых территорий с подходящими для жизни условиями и др.
На численность популяции влияют одновременно многие факторы, которые неизбежно приводят к периодическим или непериодическим, сезонным или годовым изменениям численности любого из известных видов животных и растений.
Роль в эволюции
В 1905 г. С.С. Четвериков прозорливо утверждал, что популяционные волны могут оказывать сильное влияние на интенсивность и направление естественного отбора. В самом деле, когда численность какой-либо популяции резко снижается, от нее могут остаться лишь немногие особи. Как во всякой случайной и немногочисленной выборке, частоты генов в этой “микросовокупности” особей будут иными, чем в исходной популяции. Но ведь именно эта немногочисленная группа дает начато новым усиленно размножающимся поколениям и тем самым определяет генетическую структуру популяции во время подъема численности.
В этом заключается “принцип бутылочного горлышка”; немногие особи, пережившие спад численности, как бы проникли через “узкое горлышко”, пронесли через него свойственные им генотипы и распространили их на всю популяцию. При этом некоторые, ранее присутствующие аллели могут быть безвозвратно утеряны (потому что погибли их носители), а концентрация других аллелей может резко возрасти, потому что их носители проскользнули через “горлышко”. В итоге произойдет случайное изменение частот генов и генотипов в популяциях.
Таким образом, популяционные волны сами по себе не вызывают наследственную изменчивость, а только способствуют изменению генофонда популяции.
“Волны жизни” — это своеобразный фактор — поставщик эволюционного материала, выводящий совершенно случайно и ненаправленно ряд генотипов на эволюционную арену.
Изоляция и ее значение в эволюции
Изоляция (от франц. “isolation” — отделение, разобщение) — исключение или затруднение свободного скрещивания между особями одного вида, ведущее к обособлению внутривидовых групп и новых видов.
Изоляция создается пространственными барьерами (водные преграды для сухопутных видов, участки суши для гидробионтов, возвышенности, разделяющие равнинные популяции, или равнины, разграничивающие горные популяции), территориальным разобщением в связи с расширением ареала.
Вследствие невозможности скрещивания особей из различных изолированных популяций в каждой из них возникает свое направление эволюционного процесса. Это со временем приводит к значительным отличиям в их генотипической структуре, ослаблению и даже полному прекращению обмена генами между популяциями.
Биологическая изоляция имеет три основные формы — эколого- эгологическую, морфофизиологическую и генетическую:
а) эколого-этологическая изоляция уменьшает вероятность встреч особей для скрещивания;
б) при морфофизиологической изоляции возникают препятствия для оплодотворения, связанные с различиями в строении органов размножения;
в) генетическая изоляция приводит либо к невозможности, либо к неэффективности скрещивания (снижение плодовитости, стерильность гибридов, снижение их жизнеспособности).
Генный поток (миграции) и его роль в эволюции
Миграция в эволюционном смысле означает переселение ряда особей за пределы мест обитания и как следствие — обновление генофонда другой популяции.
Скрещивание между мигрантами и особями других популяций ведет к перекомбинации генов на межпопуляционном уровне, то есть генный поток также является поставщиком материала для естественного отбора.
Причина генного потока — неполная изоляция между соседними популяциями.
Все рассмотренные нами факторы носят случайный, ненаправленный характер. Они сами по себе не могут вызвать целенаправленное изменение генофонда популяции и не могут привести к элементарному эволюционному явлению.
— Единственный фактор, имеющий направленный характер — это естественный отбор. Дрейф генов волны жизни, изоляция, миграция могут лишь повышать или понижать его эффективность.
ГРУППА 401 ХИМИЯ 21, 22
ТЕМА 21:Реакции замещения. Реакции изомеризации.
Реакции замещения.
Реакции, в результате которых осуществляется замена одного атома или группы атомов в исходной молекуле (субстрате) на другие атомы или группы атомов, называются реакциями замещения. В реакции замещения вступают предельные и ароматические соединения, такие, как, например, алканы, циклоалканы или арены.
СН4 + Сl2 → СН3Сl + НСl 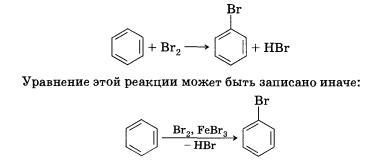
Реакции изомеризации.
Реакции, в результате которых из молекул одного вещества образуются молекулы, других веществ того же качественного и количественного состава, т. е. с той же молекулярной формулой, называют реакциями изомеризации.
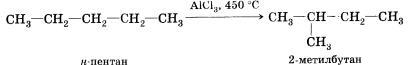
В общем, рассматривая основные принципы реагирования органических соединений, можно выделить следующие конкретные типы химических реакций:
1) Реакции замещения: галогенирование, нитрование.
2) Реакции присоединения: гидрирование, галогенирование, гидратация, гидрогалогенирование.
3) Реакции отщепления: дегидрирование, дегалогенирование, дегидратация, дегидрогалогенирование.
4) Реакции изомеризации.
Также выделяют:
5) Реакции разложения: крекинг, гидролиз, брожение.
6) Реакции полимеризации.
7) Реакции поликонденсации.
8) Реакции окисления: полное окисление (горение), неполное окисление (на катализаторе).
ТЕМА 22:Алканы. Алканы: гомологический ряд, изомерия и номенклатура алканов.
Гомологический ряд алканов
Трудно себе представить наш быт без голубых язычков огня кухонной плиты или газовой зажигалки. В этих устройствах сгорают углеводороды одного и того же класса — алканы. Различие лишь в том, что бытовой газ представляет собой главным образом метан СН4, а в резервуаре зажигалки находятся под давлением сжиженные газы пропан С3Н8 и бутан С4Н10.
Углеводороды, молекулы которых не содержат кратных связей, называют предельными или насыщенными.
Своё название предельные углеводороды получили потому, что все валентности углеродных атомов в их молекулах насыщены атомами водорода до предела.
Сокращённая структурная формула пропана СН3—СН2—СН3. Для бутана возможно написание двух структурных формул: в одной углеродная цепь имеет линейное, а в другой — разветвлённое строение: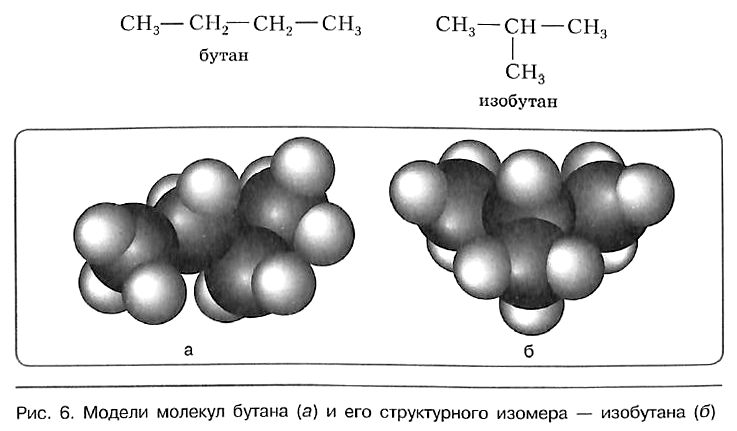
Свойства бутана и изобутана различны: бутан имеет более низкие температуры кипения и плавления. По отношению друг к другу бутан и изобутан являются изомерами, различающимися строением углеродной цепи. Изомерия углеродной цепи — вариант структурной изомерии.
| Алканы — это предельные углеводороды с общей формулой СnН2n+2, в молекулах которых все атомы соединены простыми (одинарными) связями. |

В молекулах алканов углеродные цепи открытые, т. е. незамкнутые. Но предельными могут быть и углеводороды, имеющие замкнутую углеродную цепь. Этот класс соединений называют циклоалканами. Некоторые циклоалканы, например циклогексан С6Н12, входят в состав нефти.
Простейший представитель алканов — метан СН4. Именно это соединение является родоначальником гомологического ряда алканов.
| Ряд расположенных в порядке возрастания относительных молекулярных масс веществ, сходных по строению и свойствам, но отличающихся друг от друга по составу на одну или несколько групп СН2 (гомологическую разность), называют гомологическим рядом. Вещества такого ряда называют гомологами. |
Формулу каждого последующего члена гомологического ряда легко получить из формулы предыдущего, заменив один атом водорода в молекуле на группу СН3. Так, второй гомолог ряда алканов — этан — имеет формулу СН3—СН3. Формулы и названия первых шести представителей алканов с неразветвлённой цепью атомов углерода приведены в таблице. 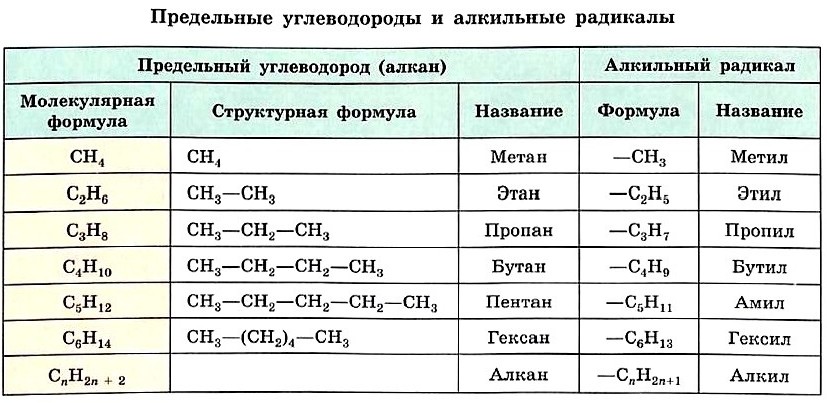
Если от молекулы алкана мысленно отщепить один атом водорода, мы получим частицу со свободной валентностью у атома углерода — радикал.
Название радикала, соответствующего предельному углеводороду, т. е. алкильного радикала, строится так: суффикс -ан заменяют на -ил, а состав будет подчиняться общей формуле СnН2n+1. Формулы и названия алкильных радикалов, приведённые в таблице, нужно запомнить, чтобы уметь называть органические вещества с разветвлённой углеродной цепью.

(с) Цитата из справочного издания «ХИМИЯ. Справочник в таблицах / М.: Издательство АЙРИС-пресс»
Номенклатура алканов
Для построения названий органических веществ, в частности предельных углеводородов, используют свод правил — номенклатуру. В соответствии с так называемой международной, или систематической, номенклатурой при составлении названий алканов руководствуются следующими правилами:
- 1) выбирают в молекуле самую длинную цепь атомов углерода;
- 2) нумеруют атомы углерода в цепи с того конца, к которому ближе разветвление;
- 3) записывают основу названия вещества — название углеводорода с тем же числом углеродных атомов, что и в пронумерованной цепи;
- 4) перед основой названия перечисляют все заместители основной цепи с указанием номеров атомов углерода, при которых они стоят. Если одинаковых заместителей несколько, перед их названиями ставят приставки ди-, три-, тетра-,
- 5) все цифры друг от друга отделяют запятыми, буквы от цифр — дефисом. Если при одном углеродном атоме имеется не один, а два заместителя, его цифра повторяется в названии дважды;
- 6) перед названием предельного углеводорода нормального (неразветвлённого) строения ставят букву н-.
Проиллюстрируем эти правила примерами названий алканов.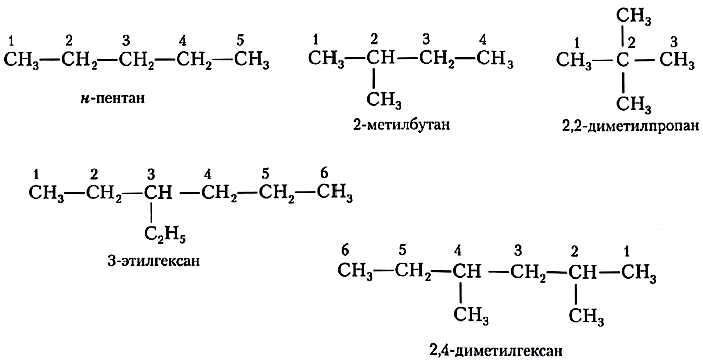
ГРУППА 401 ЭКОЛОГИЯ 22
ТЕМА 22:Законы биологической продуктивности.
Биологическая продуктивность экосистем
Продуктивность экосистем тесно связана с потоком энергии.
Биологическая продуктивность – это воспроизведение биомассы растений, микроорганизмов и животных, входящих в состав биогеоценоза.
Органическое вещество, создаваемое продуцентами в процессе фотосинтеза называется первичной продукцией экосистемы. Первичной продукцией определяется общий поток энергии через биотический компонент экосистемы и биомасса живых организмов, которые могут существовать в экосистеме. Делится на: валовую (общее количество созданного органического вещества) и чистую (оставшаяся после расходов на дыхание и корневые выделения).
Первичная продуктивность – скорость, с которой автотрофные организмы (продуценты) в процессе фотосинтеза связывают энергию и запасают ее в форме органического вещества.
Консументы образуют свою биомассу. Биомасса и скорость ее образования консументами - вторичная продукция, т.е. продукция гетеротрофных организмов.
Вторичная продуктивность - скорость образования продукции гетеротрофами.
Кроме первичной и вторичной продукции биоценозов, различают промежуточную (после потребления другими членами биогеоценоза возвращается в круговорот веществ этой же системы) и конечную (исключается из данного биогеоценоза, т.е. выводится за его пределы) продукцию.
Количественные соотношения первичной и вторичной продукции в экосистемах подчиняются правилу пирамиды.
Экологические пирамиды
Для изучения взаимоотношений между организмами в экосистеме и для их графического изображения используют экологические пирамиды.
Они выражают трофическую структуру экосистемы в геометрической форме. Основанием пирамиды служит уровень продуцентов, а последующие уровни питания образуют этажи и вершину пирамиды.
Экологические пирамиды отражают характеристики любого биоценоза: их высота пропорциональна длине рассматриваемой пищевой цепи, т.е. числу содержащихся в ней трофических уровней; их форма отражает эффективность превращения энергии при переходе с одного уровня на другой.
Основанием пирамиды служит уровень продуцентов, а последующие уровни питания образуют этажи и вершину пирамиды.
Известны три основных типа экологических пирамид:
пирамида чисел (пирамида Элтона – 1927 г.) – отражает численность организмов на каждом трофическом уровне;
2. пирамида биомассы – отражает пищевые взаимоотношения в экосистеме, в ней учитывается суммарная масса организмов (биомасса) каждого трофического уровня;
пирамида энергии – отображает связи между организмами на разных трофических уровнях.
В 1942 г. Р. Линдеман сформулировал закон пирамиды энергии (закон 10 %) – с одного трофического уровня экологической пирамиды переходит на другой ее уровень не более 10 % энергии.
В наземных экосистемах действует правило пирамиды биомасс:суммарная масса растений превышает массу всех травоядных, а их масса превышает всю биомассу хищников.
Пирамиды чисел и пирамиды биомасс не всегда имеют классический вид, только пирамиды накопленной энергии – всегда.
Основные законы и правила экологии
закон минимума Ю. Либиха: жизненные возможности организмов и экосистем определяются экологическими факторами, количество и качество которых близки к необходимому минимуму;
закон толерантности, или выносливости, В. Шелфорда: лимитирующим фактором процветания организмов (видов) может быть и максимум экологического воздействия. Диапазон между минимумом и максимумом экологического фактора определяет выносливость (толерантность) организмов к нему;
закон внутреннего динамического равновесия: вещество, энергия, информация и динамические качества отдельных природных систем в их иерархии взаимосвязаны настолько, что любое изменение одного из них вызывает сопутствующие функционально- структурные количественные и качественные перемены, где происходят такие преобразования;
закон константности количества живого вещества биосферы В.И. Вернадского: для данного геологического периода количество живого вещества биосферы - постоянная величина;
закон пирамиды энергии Р. Линдемана, или правило 10 %: с одного трофического уровня на другой в среднем переходит 10 % энергии;
закон 1 %: изменение энергетики природной системы в среднем на 1 % выводит экосистему из состояния равновесия;
закон максимума биогенной энергии Вернадского- Бауэра: биосистемы, находясь в состоянии динамического равновесия с окружающей средой и эволюционно развиваясь, увеличивают воздействие на среду;
закон необратимости эволюции Л. Полло: любой организм, популяция, вид не могут вернуться к состоянию предков;
закон сукцессионного замедления: в зрелых стабильно- равновесных экосистемах процессы замедляются;
закон равнозначности всех условий жизни: все природные условия среды, необходимые для жизни, равнозначны;
закон последовательности прохождения фаз развития: каждая природная экосистема проходит определенные эволюционные фазы от простой к сложной;
закон физико- химического единства живого вещества В.И. Вернадского: на нашей планете все живое вещество в физико- химическом отношении едино;
закон необходимого разнообразия: экосистемы не могут сформироваться из абсолютно одинаковых элементов;
правило замещения экологических условий В.В. Алехина: в определенной степени любое условие среды может быть замещено другим;
принцип неполноты (неопределенности) информации: информация, необходимая для проведения различных мероприятий по преобразованию природы, недостаточна для априорного суждения о результатах в связи со сложностью и своеобразием экосистем и непредвиденностью некоторых цепных реакций;
ГРУППА 408 ХИМИЯ 33, 34
ТЕМА 33:Аминокислоты, их строение, изомерия и свойства. Аминокислоты как амфотерные органические соединения.
ТЕМА 34:Генетическая связь аминокислот с другими классами органических соединений.
