СРЕДА 29.12.21Г. 306,403,408
|
Практическая работа №3 |
Цель работы: повторить основные качественные реакции органических веществ, научиться решать экспериментальные задачи на распознавание органических веществ.
Реактивы и оборудование: раствор KMnO4(розовый), Аммиачный раствор оксида серебра – реактив Толленса[Ag(NH3)2]OH (упрощённо +Ag2O NH3 раствор→), раствор FeCl3(светло-жёлтый), свежеосаждённый гидроксид меди (II) в сильнощелочной среде, лакмус, фенолфталеин, насыщенный раствор соли кальция.
Литература: Г.Е.Рудзитис, Ф.Г.Фельдман, химия 10 класс, стр.149
С правилами по технике безопасности ознакомлен:______________________(подпись)
Ход работы:
Оформить работу в виде отчетной таблицы.
Отчетная таблица
Что делали | Что наблюдали | Уравнения реакций | Выводы |
|
|
|
|
|
|
|
1.Распознавание уксусной кислоты (уксусная кислота –одна из самых древних кислот ,которую удалось выделить и использовать человечеству. В организме человека за сутки образуется до 400 грамм этой кислоты)
СuSО4 + 2NаОН → | Сu(ОН)2 ↓ | + Nа2SО4 |
голубой осадок |
2СН3 – СООН + Сu(ОН)2 → (СН3 – СОО)2Сu + Н2О
Получить свежеосажденный гидроксид меди. Прилить к нему 1 – 2 мл этанола. Голубой осадок растворяется, образуя раствор такого же цвета.
2. Распознавание глюкозы
СuSО4 + 2NаОН → | Сu(ОН)2 ↓ | + Nа2SО4 |
голубой осадок |
СН2ОН – (СНОН)4 – СНО + 2Сu(ОН)2 ![]() СН2ОН – (СНОН)4 – СООН + Сu2О↓ +2Н2О
СН2ОН – (СНОН)4 – СООН + Сu2О↓ +2Н2О
К 1 – 2 мл глюкозы прилить свежеосажденный гидроксид меди. Сначала осадок растворяется, затем при нагревании раствор приобретает окраску от красной до желто – оранжевой. Это свидетельствует о наличии в исследуемой жидкости глюкозы.
3. Распознавание глицерина(глицерин входит в состав мазей, в пищевой промышленности глицерин под кодом Е-422).
2СН2ОН – СНОН – СН2ОН + Сu(ОН)2 → глицерат меди (II) + 2Н2О
К 2 мл глицерина прилить свежеосажденный гидроксид меди. Раствор окрашивается в ярко – синий цвет в результате образования комплексного соединения глицерата меди (II).
4. Распознавание сахарозы. Сахар-сложное орган.в-во, содержащее много С. Чтобы доказать возьмем немного сахара и добавим к нему серную кислоту. Она забирает воду, получается свободный углерод(черное в-во) С12Н22О11-----серн.к-та------------12С+11Н2О
5.Распознавание крахмала. Капнем р-ром иода на картофель и белый хлеб. Если образуется синее пятно, то они содержат крахмал.
6. Определение кислотности р-ров глицерина и мыла с помощью имеющихся индикаторов.
(лакмус, фенолфталеин,метилоранж). Глицерин имеет слабокислую среду, а мыло-щелочную.
Общий вывод(на основе цели)!
(((
1. Распознавание уксусной кислоты (уксусная кислота – одна из самых древних кислот, которую удалось выделить и использовать человечеству. В организме человека за сутки образуется до 400 грамм этой кислоты) СuSО4 + 2NаОН →Сu(ОН)2 ↓+ Nа2SО4
2СН3 – СООН + Сu(ОН)2 → (СН3 – СОО)2Сu + Н2О
Получить свежеосажденный гидроксид меди. Прилить к нему 1 – 2 мл этанола. Голубой осадок растворяется, образуя раствор такого же цвета.
2. Распознавание глюкозы
СuSО4 + 2NаОН → Сu(ОН)2 ↓ + Nа2SО4
СН2ОН – (СНОН)4 – СНО + 2Сu(ОН)2 СН2ОН – (СНОН)4 – СООН + Сu2О↓ +2Н2О
К 1 – 2 мл глюкозы прилить свежеосажденный гидроксид меди. Сначала осадок растворяется, затем при нагревании раствор приобретает окраску от красной до желто – оранжевой. Это свидетельствует о наличии в исследуемой жидкости глюкозы.
3. Распознавание глицерина (глицерин входит в состав мазей, в пищевой промышленности глицерин под кодом Е-422).
2СН2ОН – СНОН – СН2ОН + Сu(ОН)2 → глицерат меди (II) + 2Н2О
К 2 мл глицерина прилить свежеосажденный гидроксид меди. Раствор окрашивается в ярко – синий цвет в результате образования комплексного соединения глицерата меди (II).
4. Распознавание сахарозы. Сахар-сложное орган. в-во, содержащее много С. Чтобы доказать возьмем немного сахара и добавим к нему серную кислоту. Она забирает воду, получается свободный углерод (черное в-во) С12Н22О11---серн.к-та--12С+11Н2О
5. Распознавание крахмала. Капнем р-ром иода на картофель и белый хлеб. Если образуется синее пятно, то они содержат крахмал.
6. Определение кислотности р-ров глицерина и мыла с помощью имеющихся индикаторов.
(лакмус, фенолфталеин,метилоранж). Глицерин имеет слабокислую среду, а мыло-щелочную.)))_
ТЕМА:
.Практическая работа №6. Распознавание пластмасс и волокон.
Тема: «Распознавание пластмасс и волокон»
Оборудование и реактивы: образцы пластмасс и волокон под номерами, спиртовка, спички, стеклянные палочки, тигельные щипцы, асбестовые сетки.
Распознавание пластмасс
В разных пакетах под номерами имеются образцы пластмасс. Пользуясь при веденными ниже данными, определите, под каким номером какая пластмасса находится.
Полиэтилен. Полупрозрачный, эластичный, жирный на ощупь материал. При нагревании размягчается, из расплава можно вытянуть нити. Горит синеватым пламенем, распространяя запах расплавленного парафина, продолжает гореть вне пламени.
Поливинилхлорид. Эластичный или жесткий материал, при нагревании быстро размягчается, разлагается с выделением хлороводорода. Горит коптящим пламенем, вне пламени не горит.
Полистирол. Может быть прозрачным и непрозрачным, часто хрупок. При нагревании размягчается, из расплава легко вытянуть нити. Горит коптящим пламенем, распространяя запах стирола, продолжает гореть вне пламени.
Полиметилметакрилат. Обычно прозрачен, может иметь различную окраску. При нагревании размягчается, нити не вытягиваются. Горит желтоватым пламенем с синей каймой и характерным потрескиванием, распространяя эфирный запах.
Фенолформальдегидная пластмасса. Темных тонов (от коричневого до черного). При нагревании разлагается. Загорается с трудом, распространяя запах фенола, вне пламени постепенно гаснет.
Распознавание волокон
В разных пакетах под номерами содержатся образцы волокон. Пользуясь приведенными ниже данными, определите, под каким номером какое волокно находится.
Хлопок. Горит быстро, распространяя запах жженой бумаги, после сгорания остается серый пепел.
Шерсть, натуральный шелк. Горит медленно, с запахом жженых перьев, после сгорания образуется черный шарик, при растирании превращающийся в порошок.
Ацетатное волокно. Горит быстро, образуя нехрупкий, спекшийся шарик темно-бурого цвета. В отличие от других волокон растворяется в ацетоне.
Капрон. При нагревании размягчается, затем плавится, из расплава можно вытянуть нити. Горит, распространяя неприятный запах.
Лавсан. При нагревании плавится, из расплава можно вытянуть нити. Горит коптящим пламенем с образованием темного блестящего шарика.
Краткие теоретические и учебно-методические материалы
по теме практического занятия
Полимерами называют вещества, молекулы которых состоят из множества повторяющихся структурных звеньев, соединенных между собой химическими связями. Существует два основных способа получения полимеров — реакции полимеризации и реакции поликонденсации.
Реакция полимеризации — это химический процесс соединения множества исходных молекул низкомолекулярного вещества (мономера) в крупные молекулы (макромолекулы ) полимера.
В реакцию полимеризации могут вступать соединения, содержащие кратные связи, то есть непредельные соединения. Это могут быть молекулы одного мономера или разных мономеров.
В первом случае происходит реакция гомополимеризации — соединение молекул одного мономера, во втором — реакция сополимеризации — соединение молекул двух и более исходных веществ.
К реакциям гомополимеризации относятся реакции получения полиэтилена, полипропилена, поливинилхлорида и т. д., например:
Выражение в скобках называют структурным звеном, а число n-в формуле полимера — степенью полимеризации.
К реакциям сополимеризации относится, например, реакция получения бутадиен-стирольного каучука.
Реакция поликонденсации — это химический процесс соединения исходных молекул мономера в макромолекулы полимера, идущий с образованием побочного низкомолекулярного продукта (чаще всего воды).
В реакции поликонденсации вступают молекулы мономеров с функциональными группами.
Например, реакция получения фенолформальдегидных смол: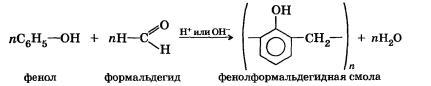
С помощью реакций поликонденсации получают полиэфиры, полиамиды, полиуретаны, полиакрил и т. д.
Пластмассы.
Пластмассами называют материалы, изготовляемые на основе полимеров. Пластмассы сочетают в себе разнообразные ценные качества, такие как лёгкость, прочность, химическая
стойкость и др., которые обусловили проникновение их в различные отрасли народного хозяйства. Кроме полимеров (их часто называют смолой) в пластмассах почти всегда содержатся другие компоненты, придающие материалу определённые качества. Полимерное вещество является для них связующим.
В пластмассы входят наполнители (древесная мука, ткань, асбест, стекловата и др.), которые улучшают их механические свойства.
Пластификаторы – повышают эластичность, устраняют хрупкость.
Стабилизаторы – способствуют сохранению свойств пластмасс в процессе их переработки и использования; красители придают необходимую окраску.
Обычные способы получения полимеров – это реакции полимеризации, лежащие в основе получения термопластичных пластмасс, и реакции поликонденсации, лежащие в основе получения термореактивных пластмасс.
Термопластичные полимеры при нагревании размягчаются и в этом состоянии легко изменяют форму, которую сохраняют при охлаждении. При следующем нагревании они снова размягчаются и могут принимать новую форму.
Термореактивные полимеры при нагревании сначала становятся пластичными, при дальнейшем нагревании утрачивают пластичность, становятся неплавкими. Повторно переработать такой полимер в новое изделие невозможно.
Наиболее типичными способами получения изделий из термопластичных пластмасс является литьё под давлением и экструзия (выдавливание), а из термореактивных пластмасс – горячее прессование.
Краткая характеристика некоторых пластмасс
Полиэтилен – твёрдый, жирный на ощупь, белого цвета термопластичный полимер. Стоек по отношению к агрессивным средам. Благодаря высокой температуре плавления, обладает существенными преимуществами перед другими материалами (полиэтиленом, полиметилметакрилатом, поливинилхлоридом), близким по свойствам.
Полипропилен идёт на изготовление высокопрочной изоляции, труб, деталей машин, химической аппаратуры. Благодаря высокой механической прочности, его используют для изготовления канатов, сетей, технических тканей.
Поливинилхлорид – обладает большой химической стойкостью, хорошими электроизоляционными свойствами и большой механической прочностью. Термопластичный полимер, на его основе изготавливают два вида пластмасс: винипласт, обладающий значительной жесткостью и пластикат – более мягкий материал.
Винипласт идёт на изготовление химически стойкой аппаратуры, ванн для никелирования, жестких плёнок. Пластикат используется для изоляции, для производства предметов широкого потребления (плащей, сумок, линолеума, клеенок, для получения материалов, заменяющих кожу – в производстве обуви).
Полиметилметакрилат – за свою прозрачность называется органическим стеклом. Обладает удовлетворительной прочностью и значительно меньшей хрупкостью, чем обычное силикатное стекло, способностью пропускать ультрафиолетовые лучи. Термопластичный полимер, находит применение в строительстве, в часовом деле, различных отраслях промышленности и в быту.
Фенолформальдегидная смола – обычно используется в смеси с наполнителями, красителями и т.п., а затем уже производят формование изделий способом горячего прессования. Термореактивный полимер. Введение различных наполнителей позволяет получить материалы, имеющие ценные свойства. Так текстолит и стеклотекстолит, армированные текстильными тканями и стеклотканью, по прочности близки к дюралюминию и стали.
Текстолит – хлопчатобумажная ткань, пропитанная фенолформальдегидной смолой и спрессованная при повышенной температуре. Устойчив к нагрузкам. Легко поддаётся механической обработке. Применяется для изготовления шарикоподшипников, шестерёнки для машин, предусмотренных для больших нагрузках.
Таблица 1 « Распознавание пластмасс»
Название пластмассы | Отношение к нагреванию | Характер горения |
Полиэтилен | Размягчается – можно вытянуть нить. | Горит синеватым пламенем, распространяя слабый запах горящего парафина. При горении отделяются капли. Вне пламени продолжает гореть. |
Поливинилхлорид (полихлорвинил) | Размягчается при 60-70˚С, выше 110-120˚С разлагается. | Горит коптящим пламенем. Вне пламени не горит. |
Полистирол | Размягчается – легко вытягиваются нити. | Горит коптящим пламенем, распространяя специфический запах. Вне пламени продолжает гореть. |
Полиметилметакрилат (орг. стекло) | Размягчается. | Горит жёлтым пламенем, с синей каймой у краев, с характерным потрескиванием, распространяя резкий запах. |
Целлулоид | Разлагается. | Горит очень быстро, оставляя следы золы. |
Фенолформальдегидные пластмасы | Разлагается при сильном нагревании | Загорается с трудом, при горении обугливается, распространяя резкий запах фенола. Вне пламени постепенно гаснет, не размягчается. |
Волокна
– природные или искусственные высокомолекулярные вещества, отличающиеся от других полимеров более высокой степенью упорядоченности молекул и, как следствие, особыми физическими свойствами, позволяющими использовать их для получения нитей. Волокна делят на натуральные (природные) и химические. Натуральные волокна могут быть растительного или животного происхождения. Химические волокна в свою очередь подразделяют на искусственные и синтетические.
Природные волокна:
Волокно растительного происхождения – хлопок, лен.
Хлопковое волокно получают из субтропического растения – хлопчатника. Хлопковое волокно легкое, достаточно прочное, мягкое, гигроскопичное.
Волокна животного происхождения – шерсть и шелк.
Шелк вырабатывают многочисленные гусеницы и пауки.
Шерсть – волокна волосяного покрова овец, коз, верблюдов и других животных.
Искусственные волокна:
Наибольшее значение среди искусственных волокон занимают ацетатное и вискозное волокна, получаемые из древесной целлюлозы.
Синтетические волокна:
из синтетических волокон наибольший интерес представляют полиамидное волокно – полиамид – 6 (капрон) и полиэфирное – полиэтиленгликольтерефталат (лавсан).
Капрон получают из капролактала, который под воздействием воды размыкает цикл, образуя ε – капроновую кислоту. Из этой кислоты в результате поликонденсации образуется полимер линейной структуры:
n H2N – (CH2)5 – COOH → [- NH – (CH2)5 – CO -]n + (n-1)H2O
Лавсан (полиэтилентерефталат) - представитель полиэфиров:

Получают реакцией поликонденсации терефталевой кислоты и этиленгликоля:
HOOC-C6H4-COOH + HO-CH2CH2-OH + HOOC-C6H4-COOH + … →
→ HOOC-C6H4-CO – O-CH2CH2-O – OC-C6H4-CO – … + nH2O
полимер-смола
В общем виде:
n HOOC-C6H4-COOH + n HO-CH2CH2-OH →
→ HO-(-CO-C6H4-CO-O-CH2CH2-O-)n-H + (n-1) H2O
Таблица 2. «Распознавание волокон»
Волокно | Сжигание |
Хлопок | Горит быстро с запахом жжёной бумаги. После горения остается серый пепел. |
Шерсть | Горит медленно с запахом жжёных перьев. После горения образуется хрупкий чёрный шарик, растирающийся в порошок. |
Ацетатное волокно | Горит быстро, образуя нехрупкий спёкшийся тёмно-бурый шарик. Вне пламени горение постепенно прекращается. |
Капрон | Плавится, образуя твёрдый блестящий шарик тёмного цвета. При горении распространяется неприятный запах. |
Лавсан | Плавится, затем горит коптящим пламенем с образованием тёмного твердого блестящего шарика. |
Нитрон | Горит, образуя тёмный рыхлый неблестящий шарик. |
Вопросы для закрепления теоретического материала к практическому занятию:
Дайте определения следующим понятиям: полимеры, структурное звено, степень полимеризации.
В чем разница между реакциями полимеризации и поликонденсации?
Какие полимеры называются термопластичными?
Какие полимеры называются термореактивными?
Дайте классификацию волокон.
Этап | Учёные | Вклад в развитие эволюционных представлений |
Зарождение эволюционных взглядов | Древние философы | Высказывали идеи естественного развития живой природы |
| Метафизический | Карл Линней ( | 1. Описал более 2. Ввёл бинарную номенклатуру. 3. Предложил первую классификацию живой природы, основанную на сравнении отдельных признаков. Ошибки: 1. считал виды неизменными, созданными творцом, а приспособленность — изначально целесообразной. 2. Классификация оказалась искусственной, так как исследователь брал во внимание 3. Не учитывал родство организмов. К концу жизни признал возможность изменения видов |
| Первое эволюционное учение | Жан Батист Ламарк ( | 1. Создал первое эволюционное учение. 2. Понимал эволюцию как процесс постепенного усложнения организации от низших организмов к высшим. Ввёл понятие градации — постепенного развития от простого к сложному. 3. Считал, что виды находятся в постоянном движении — постепенно изменяются и превращаются в другие виды. Существуют только переходные формы. 4. Показал приспособительный характер изменений. 5. Впервые разделил животных на две большие группы: беспозвоночных и позвоночных. Ошибки: 1. неверно объяснял причины и движущие силы эволюции. Утверждал, что к изменениям приводит внутреннее стремление организмов к усовершенствованию и способность организмов целесообразно реагировать на изменение условий. 2. Считал, что признаки могут изменяться в результате «упражнения и неупражнения» органов. 3. Утверждал, что полезные признаки обязательно передаются потомкам. 4. Отрицал реальность существования видов |
| Теория естественного отбора | Чарльз Дарвин ( | Создал учение о естественном отборе |
Теоретический материал для самостоятельного изучения
В исторической хронологии развития эволюционных идей в биологии выделяется четыре этапа.
Первый этап - появляются предпосылки к становлению эволюционной биологии, выделению основных направлений изучения эволюции (появляются такие теории, как самозарождение жизни, влияние условий среды на организмы). Основными мыслителями того времени являются Аристотель, Демокрит, Эмпедокл.
На втором этапе основной теорией происхождения и развития жизни на Земле была теологическая, божественная.
Третий этап характеризуется началом классификации живых организмов, где основным направлением становится систематика, выдвижением глобальных торий эволюции организмов и открытием новых законов. Основными мыслителями этого времени являются: Ламарк, Кювье, Бюффон, Линней, Дарвин, Мендель.
Жорж Луи Бюффон высказывал прогрессивные идеи об изменяемости видов под влиянием условий среды (климата, питания и т.д.), выдвинул теорию, о том, что человек произошел от обезьяны.
Жорж Кювье стал основателем сравнительной анатомии. Его теория выявляет взаимосвязь всех органов и структур между собой, а их строение и функционал ставит в зависимость от условий окружающей среды, питания, размножения.
Ламарк, продолжая идею Линнея о классификации организмов, выдвинул законы, из которых следует, что у живых организмов развиваются те органы, которые им более необходимы, а приобретенные улучшения наследуются.
Чарльз Дарвин, строя теорию эволюции, утверждал, что окружающий мир постоянно меняется, а уменьшение ресурсов и ограниченность доступа к ним приводит к борьбе за выживание, в результате остаются самые сильные организмы, которые способны дать сильное потомство - происходит естественный отбор.
На четвертом этапе для изучения теории эволюции предлагается синтетическая теория, включающая в себя положения теории эволюции Дарвина (естественный отбор) и генетику (механизм формирования вида).
При изменении условий окружающей среды организму необходимо к ним приспособиться, что провоцирует изменения на генном уровне. В процессе приспособления организмов к окружающей среде выживают наиболее устойчивые к условиям среды, наименее устойчивые погибают, происходит естественный отбор. Среди организмов идет как конкуренция, так и взаимопомощь (у некоторых видов).
Тема: Чарльз Дарвин и его теория происхождения видов.
История развития и происхождения человеческого вида на протяжении столетий волнует учёных и множество обычных людей. Во все времена на этот счёт выдвигались всевозможные теории.
РОДИВШИЙСЯ В СЕМЬЕ ВРАЧА, ЧАРЛЬЗ ДАРВИН, ОБУЧАЯСЬ В КЕМБРИДЖЕ И ЭДИНБУРГЕ, СТАЛ ОБЛАДАТЬ ГЛУБОКИМИ ПОЗНАНИЯМИ В ОБЛАСТИ ГЕОЛОГИИ, БОТАНИКИ И ЗООЛОГИИ, А ТАКЖЕ ОВЛАДЕЛ НАВЫКАМИ ПОЛЕВЫХ ИССЛЕДОВАНИЙ, К КОТОРЫМ ИСПЫТЫВАЛ ТЯГУ.
Огромное влияние на формирование мировоззрения Дарвина, как учёного, оказал труд «Принципы геологии» Чарльза Лайеля – английского геолога. Согласно его мнению, современный вид нашей планеты постепенно формировался под воздействием тех же естественных сил, которые продолжают воздействовать и в наши дни. Чарльз Дарвин, естественно, был знаком с идеями Жана Батиста Ламарка, Эразма Дарвина и некоторых других эволюционистов раннего периода, но ни одна из них не подействовала на него, как теория Лайлея.
Однако поистине судьбоносную роль в судьбе Дарвина сыграло его странствие на судне «Бигль», проходившее с 1832 по 1837 годы. Сам Дарвин говорил, что самое больше впечатление на него произвели следующие открытия:
- Открытие ископаемых животных гигантских размеров и покрытых панцирем, который был похож на панцирь знакомых всем нам броненосцев;
- Очевидность того, что виды животных, близких по роду, замещают друг друга по мере продвижения по южноамериканскому материку;
- Очевидность того, что виды животных на различных островах Галапагосского архипелага лишь немногими отличаются друг от друга.
Впоследствии учёный заключил, что вышеназванные факты, как и множество других, могут быть объяснены лишь если предположить, что каждый из видов претерпевал постоянные изменения.
После того как Дарвин вернулся из путешествия, он занялся обдумыванием проблемы происхождения видов. Идей рассматривалось множество, в том числе, и идея Ламарка, но все они были отброшены за отсутствием объяснений поразительной способности растений и животных приспосабливаться к условиям среды обитания. Этот факт, который ранние эволюционисты считали не требующим обоснований, стал для Дарвина наиболее важным вопросом. Так он начал собирать информацию на тему изменчивости растений и животных в природных и домашних условиях.
Много лет спустя, вспоминая о возникновении своей теории, Дарвин писал, что совсем скоро он осознал, что главное значение в успешном создании человеком полезных видов растений и животных имел именно отбор. Хотя, на протяжении определённого времени учёный всё ещё не мог понять, как отбор может быть применимым к тем организмам, которые живут в естественной среде.
Именно в этот период в научных кругах Англии активно обсуждались идеи Томаса Мальтуса – английского учёного и демографа, который говорил о том, что популяция населения растёт в геометрической прогрессии. После прочтения его работы «О народонаселении» Дарвин продолжил свою предыдущую мысль, сказав, что продолжительные наблюдения за образом жизни растений и животных подготовили его к оценке значения вездесущей борьбы за существование. Но он был поражён мыслью, что благоприятные изменения в таких условиях должны оставаться и сохраняться, а неблагоприятные должны быть подвергнуты уничтожению. Итогом всего этого процесса и должно быть появление новых видов.
В результате, в 1838 году у Дарвина появилась теория происхождения видов посредством естественного отбора. Однако публикация этой теории состоялась лишь в 1859 году. А поводом для публикации послужили довольно-таки драматические обстоятельства.
В 1858 году человек по имени Альфред Уоллес – молодой британский биолог, натуралист и путешественник прислал Дарвину рукопись своей статьи «О тенденции разновидностей к неограниченному отклонению от первоначального типа». В данной статье было представлено изложение теории происхождения видов через естественный отбор. Дарвин решил не публиковать свой труд, но его соратники Чарльз Лайель и Джозеф Долтон Гукер, уже давно знавшие об идеях своего товарища и знакомые с набросками его работы, смогли убедить Дарвина, что публикация работы должна состояться одновременно с публикацией работы Уоллеса.
Так, в 1959 году вышла в свет работа Чарльза Дарвина «Происхождение видов путём естественного отбора, или Сохранение благоприятных рас в борьбе за жизнь», и её успех был просто ошеломительным. Теория Дарвина была прекрасно принята и поддержана одними учёными и жёстко раскритикована другими. Но все последующие труды Дарвина, как и этот, после публикации мгновенно приобретали статус бестселлеров и издавались на многих языках. Сам же учёный в мгновение ока приобрёл мировую известность.
И одной из причин популярности теории Дарвина явились её базовые принципы.
ГЛАВНЫЕ ПРИНЦИПЫ ТЕОРИИ ПРОИСХОЖДЕНИЯ ВИДОВ ЧАРЛЬЗА ДАРВИНА
Вся суть теории происхождения видов Дарвина заключается в комплексе положений, являющихся логичными, способных быть проверенными экспериментально и подтверждённых фактами. Этими положениями являются следующие:
- Любой вид живых организмов включает в себя огромнейший спектр индивидуальной генетической изменчивости, которая может различаться морфологическими, физиологическими, поведенческими и любыми другими признаками. Эта изменчивость может быть непрерывного количественного или прерывистого качественного характера, однако существует в любой момент времени. Найти двух особей, которые были бы абсолютно идентичны по совокупности признаков, невозможно.
- У любого живого организма есть способность быстро увеличивать популяцию. Не может быть никакого исключения из правила, говорящего о том, что органические существа размножаются в такой прогрессии, что если бы они не истреблялись, то одна пара могла бы покрыть потомством всю планету.
- Для любого вида животных имеются только ограниченные ресурсы для жизни. По этой причине большое производство особей должно служить катализатором борьбы за существование либо между представителями одного вида, либо между представителями различных видов, либо с условиями существования. Борьба за существование, согласно теории Дарвина, включает в себя как борьбу представителя какого-либо вида за жизнь, так и его борьбу за успешное обеспечение себя потомством.
- В борьбе за существование способны выжить и успешно производить потомство только самые приспособленные особи, которые имеют особые отклонения, оказавшиеся адаптивными к конкретным условиям среды обитания. Причём, такие отклонения возникают именно случайно, а не реагируя на воздействие среды. И полезность этих отклонений также случайна. Отклонение передаётся потомкам особи, которая выжила, на генетическом уровне, по причине чего они становятся более адаптированными к имеющейся среде обитания, нежели другие особи того же вида.
- Естественный отбор представляет собой процесс выживания и преимущественного размножения приспособленных представителей популяции. Естественный отбор, по словам Дарвина, точно так же постоянно фиксирует любые изменения, сохраняет хорошие и отбрасывает плохие, как это делает селекционер, изучающий множество особей и отбирающий и размножающий лучше из них.
- Применительно к отдельным изолированным разновидностям в различных условиях жизни, естественный отбор приводит к расхождению их признаков и, в итоге, к образованию нового вида.
Данные положения, являющиеся практически безупречными с точки зрения логики, и подкреплённые большим количеством фактического материала, являются основой теории эволюции, которую мы можем видеть сегодня.
ГРУППА 408 БИОЛОГИЯ 22
ТЕМА: Регуляция работы генов у прокариот и эукариот.
Регуляция активности генов. Опероны у бактерий
Все клетки любого организма, какие бы функции они ни выполняли, имеют полный набор свойственных данному организму генов. Вместе с тем хорошо известно, что у любого организма клетки разных тканей и органов отличаются поразличным признакам и набору имеющихся в них белков. Даже в одной клетке на разных стадиях ее развития синтезируются и функционируют разные белки. Следовательно, располагая полной генетической информацией, каждая клетка на определенном этапе развития использует лишь ту ее часть, которая необходима в настоящий момент, транскрибируются только те гены, продукты которых нужны клетке в данный момент для осуществления ее функций. Следовательно, клетка должна располагать механизмами, определяющими, какие гены и в какой последовательности должны экспрессироваться ("выражаться", то есть давать продукт — РНК или белок). Наиболее полно регуляция генной активности изучена на примерах адаптивного синтеза ферментов у прокариот.
В зависимости от условий количество определенного фермента в бактериальной клетке может существенно изменяться. Некоторые ферменты, необходимые бактерии для усвоения определенных питательных веществ, активно синтезируются в клетке только тогда, когда эти вещества присутствуют в культурной среде, и синтез их прекращается, если каким-либо образом они удаляются из среды. Такой тип регуляции синтеза фермента называется индукцией, а вещество, включающее экспрессию гена — индуктором.
Активация и репрессия оперонов у бактерий
лактозный оперон
Один из наиболее наглядных примеров данного типа регуляции — лактозный оперон кишечной палочки — группа генов, контролирующая синтез ферментов, осуществляющих катаболизм (расщепление) молочного сахара — лактозы. Буквально через несколько минут после добавления в питательную среду для кишечной палочки лактозы бактерии начинают вырабатывать три фермента: галактозидпермеазу, бета-галактозидазу и галактозидтрансацетилазу. Как только ресурсы лактозы в среде исчерпываются, синтез ферментов сразу же прекращается.
Приведенный пример станет более понятным при рассмотрении схемы работы лактозного оперона, изучение которого позволило французским ученым Ф. Жакобу и Ж. Моно разработать собственно концепцию оперона и выяснить основные принципы регуляции транскрипции у прокариотов.
Оперон — это группа генов прокариот, находящихся под общим промотором. Все эти гены транскрибируются на одну общую молекулу мРНК. Такая мРНК, содержащая информацию о нескольких белках, называется полицистронной. Участок ДНК или РНК, содержащий информацию об одном белке, называется цистроном.
Лактозный оперон начинается с регуляторного участка, предназначенного для присоединения белка-активатора, в свою очередь необходимого для присоединения к следующему за этим участком промотору (П) РНК-полимеразы. Последовательность нуклеотидов промотора узнаётся РНК-полимеразой.
С промотором перекрывается следующий участок — оператор (О). С ним может связываться регуляторный белок-репрессор. Репрессор блокирует промотор и тем самым предотвращает транскрипцию гена.
За оператором следуют структурные гены для трех упомянутых ранее ферментов. Заканчивается оперон терминатором, прекращающим продвижение РНК-полимеразы и транскрипцию оперона.
Регуляторный белок-репрессор в незначительном количестве синтезируется в клетке постоянно, так что в цитоплазме одновременно присутствует не более 10 его молекул. Этот белок обладает сродством к последовательности нуклеотидов в области оператора и сродством к лактозе.
В отсутствие лактозы белок-репрессор связывается с операторным участком и препятствует продвижению по ДНК РНК-полимеразы: не синтезируется мРНК, не синтезируются и ферменты. После добавления в среду лактозы белок-репрессор связывается с нею быстрее, чем с операторным участком. В результате последний остается свободным и не препятствует продвижению РНК-полимеразы. Идет транскрипция и трансляция. Синтезирующиеся ферменты осуществляют транспорт в клетку и расщепление лактозы. После того как вся лактоза будет израсходована, нечем станет связывать белок-реп рессор и он снова свяжется с оператором, прекратив транскрипцию оперона. Таким образом, индукция оперона вызывается тем, что регуляторный белок не прикрепляется к оператору. Такой тип индукции называется негативным.
Параллельно наблюдается и другой тип регуляции — позитивная регуляция. При глюкозном голодании в клетке из АТФ образуется сигнальное вещество цАМФ, которое связывается с белком-активатором (САР), после чего последний приобретает способность связывать ДНК в промоторной области и усиливать транскрипцию лактозного оперона. Таким образом, когда не хватает глюкозы, стимулируется всасывание и катаболизм лактозы. При одновременном присутствии глюкозы и лактозы последняя не метаболизируется, пока существенно не упадёт концентрация глюкозы.
триптофановый оперон
В случае индукции лактозного оперона аллолактоза (индуктор) препятствует присоединению белка-репрессора к оператору, то возможен и другой вариант регуляции, когда, наоборот, индуктор придает регуляторному белку способность присоединяться к оператору. Если в первом случае соединение индуктора с белком-регулятором разрешало транскрипцию, то во втором оно запрещает ее.
Примером такой регуляции может служить хорошо изученный триптофановый оперон кишечной палочки. В его состав входят пять структурных генов, обеспечивающих синтез аминокислоты триптофана, оператор и промотор. Репрессор синтезируется вне триптофанового оперона. Пока клетка успевает расходовать весь синтезирующийся триптофан, оперон работает, синтез триптофана продолжается. Если же в клетке появляется избыток триптофана, он соединяется с репрессором и изменяет его таким образом, что этот белок связывается с оператором. Комплекс репрессора с триптофаном взаимодействует с оператором и препятствует транскрипции структурных генов, вследствие чего синтез триптофана прекращается. В отсутствие триптофана репрессор лишается способности связываться с оператором, и происходит транскрипция структурных генов оперона и, в итоге, синтез триптофана в клетке.
Другой уровень регуляции триптофанового оперона включает аттенуацию — тонкую подстройку количества продукта в зависимости от концентрации присутствующего триптофана.
регуляция генов бактериофагов
Описанные типы регуляций характеризуют механизмы регуляции отдельных оперонов, практически не касаясь регуляции экспрессии генома в целом, в то время как совершенно очевидно, что регуляция разных оперонов должна носить согласованный характер. Такой согласованный характер работы разных оперонов и генов получил у вирусов и фагов название каскадной регуляции. Согласно принципу каскадной регуляции сначала происходит транскрипция «предранних», затем «ранних» и наконец «поздних» генов в зависимости от того, какие белки требуются на разных стадиях вирусной (фаговой) инфекции.
Конечно, принцип каскадной регуляции у фагов относится к наиболее простым. У более сложно организованных организмов для осуществления большого количества функций, происходящих одновременно или с определенной последовательностью, необходима согласованная работа многих генов и оперонов. Особенно это касается эукариотов, отличающихся не только более сложной организацией генома, но и многими другими особенностями механизмов регуляции генной активности.
регуляция генов эукариот
По принципам регуляции гены эукариотов можно условно разделить на три группы: 1) функционирующие во всех клетках организма; 2) функционирующие только в тканях одного типа; 3) обеспечивающие выполнение специализированными клетками конкретных функций.
Механизмы регуляции экспрессии генов у эукариот:
- У эукариот известна регуляция генной активности на уровне структуры хроматина. В регуляции генов эукариот важную роль играют гистоны — основные белки, входящие в состав хромосом. Одни модификации гистонов характерны для активно работающих генов, другие — для молчащих. Существуют также вариантные гистоны, закодированные особыми генами. Они могут заменять "обычные" гистоны в определенных хроматиновых контекстах, влияя на регуляцию генов.
На электронных микрофотографиях в ядрах неделящихся клеток эукариот видны более плотно упакованные участки — гетерохроматин — и более рыхлые участки, называемые эухроматином. Гетерохроматин содержит центромерные и теломерные участки хромосом (облигатный гетерохроматин) и молчащие гены (факультативный гетерохроматин), а эухроматин — активно работающие гены. - Также распространенным типом регуляции экспрессии генов у эукариот является метилирование ДНК, в основном по 5 положению цитозина. Метилированная ДНК, как правило, присутствует в выключенных генах. Этим, в частности, объясняется трудность организменного клонирования, связанная с тем, что в соматических клетках (клетках тела) многие гены метилированы, поэтому когда ядром соматической клетки замещают ядро зиготы, экспрессия этих генов зачастую не активируется, т.к. метилирование генов не снимается. Надо отметить, что регуляторное метилирование распространено не у всех эукариот, например, оно не характерно для генома мушки дрозофилы.
- Еще одним существенным отличием транскрипции у эукариотов является то, что многие мРНК длительное время сохраняются в клетке в виде особых частиц— информосом, в то время как мРНК прокариотов практически еще в процессе транскрипции поступают в рибосомы, транслируются, после чего быстро разрушаются. У эукариот развита регуляция экспрессии на уровне стабильности (времени жизни), трансляционной активности, локализации мРНК.
- У эукариот известны и другие типы регуляции активности генов, такие, как эффект положения или дозовая компенсация. В первом случае речь идет об изменении генной активности в зависимости от конкретного окружения: перемещение гена из одного места хромосомы в другое может приводить к изменению активности как этого гена, так и близлежащих. Во втором случае нехватка одной дозы какого-либо гена (в первую очередь это относится к генам, локализованным в половых хромосомах гетерогаметного пола, когда одна из гомологичных половых хромосом либо генетически инертна, либо полностью отсутствует) фенотипически не проявляется за счет компенсаторного увеличения активности оставшегося гена. В целом же регуляция активности генов у эукариот в настоящее время активно изучается.
- Вместе с тем имеется много данных, указывающие, что транскрипция определенных генов эукариот также может осуществляться скоординированно. Энхансеры — активирующие элементы в ДНК эукариот — могут действовать на огромных расстояниях, в десятки и сотни тысяч пар нуклеотидов. Предполагается, что это происходит путем образования хроматиновых петелль. Энхансер может действовать на целую группу генов; распространение действия энхансеров блокируется элементами-инсуляторами. Возможно, один энхансер активирует один петлевой домен ДНК — участок ДНК, формирующий одну хромомерную петлю и содержащий совместно регулирующиеся гены. Однако далеко не всегда совместно регулирующиеся гены у эукариот расположены рядом; трудно доказать и гипотезу петлевых доменов в связи со сложностями их картирования.

Комментариев нет:
Отправить комментарий