07.12.22 г. СРЕДА . Группы 403,508, 306,408
Здравствуйте, уважаемые студенты, записывайте дату, тему и выполняйте необходимые записи (ВСЁ подряд не пишите, читайте, выбирайте, можно составить план, ЕСЛИ ЕСТЬ ВИДЕО, НАДО ПОСМОТРЕТЬ ,ВЫПОЛНИТЬ ПО НЕМУ ЗАПИСИ, МНОГО НЕ НУЖНО ПИСАТЬ. Материала может быть выложено много, но это не значит, что всё надо записывать! После этого, сфотографируйте и отошлите мне на почту rimma.lu@gmail.com . Тетрадь привезете, когда перейдем на очную форму обучения).Справа находится АХИВ БЛОГА , смотрите дату и номер своей группы
моя почта : rimma.lu@gmail.com Жду ваши фотоотчеты!
ГРУППА 403 экология 3,4
ТЕМА 3,4 : ОБЩИЕ ЗАКОНЫ ЗАВИСИМОСТИ ОРГАНИЗМОВ ОТ ФАКТОРОВ СРЕДЫ.
ОТКРОЙТЕ ЭЛЕКТРОННЫЙ УЧЕБНИК С.15 ПАРАГРАФ 2, ПРОЧИТАЙТЕ, РАССМОТРИТЕ РИСУНКИ, СХЕМЫ, СОСТАВЬТЕ ПЛАН, НА С.21 ВОПРОСЫ, ОТВЕТЬТЕ ПИСЬМЕННО НА ВОПРОС 1.
ГРУППА 508 БИОЛОГИЯ 14,15
ТЕМА 14,15 : Обмен веществ.Фотосинтез.
ПРОЧИТАЙТЕ, РАССМОТРИТЕ РИСУНКИ, СОЗДАЙТЕ КРАТКИЙ КОНСПЕКТ,МОЖНО ЧЕРТИТЬ СХЕМЫ,ГДЕ ЭТО НАДО.
Обязательным условием существования любого организма является постоянный приток питательных веществ и постоянное выделение конечных продуктов химических реакций, происходящих в клетках. Клетка постоянно находится в движении – цитоплазма перемещается, увлекая за собой органоиды и включения, активно работают рибосомы и митохондрии, совершается множество химических превращений. Все живые организмы, существующие на Земле, представляют собой открытые системы, характеризующиеся способностью активно обмениваться с окружающей средой веществами и энергией. Из окружающей среды в клетку поступают различные вещества, а из клетки в окружающую среду удаляются ненужные продукты обмена – происходит обмен веществ, или метаболизм (Рис. 1).

Рис. 1. Обмен веществ клетки с окружающей средой (Источник)
Питательные вещества используются организмами в качестве источника атомов химических элементов (прежде всего атомов углерода), из которых строятся либо обновляются все структуры. В организм, кроме питательных веществ, поступают также вода, кислород, минеральные соли.
Поступившие в клетки органические вещества (или синтезированные в ходе фотосинтеза) расщепляются на строительные блоки – мономеры и направляются во все клетки организма (Рис. 2). Часть молекул этих веществ расходуется на синтез специфических органических веществ, присущих данному организму. В клетках синтезируются белки, липиды, углеводы, нуклеиновые кислоты и другие вещества, которые выполняют различные функции (строительную, каталитическую, регуляторную, защитную и так далее).
Другая часть низкомолекулярных органических соединений, поступивших в клетки, идет на образование АТФ, в молекулах которой заключена энергия, предназначенная непосредственно для выполнения работы.

Рис. 2. Распределение органических веществ
Энергия необходима для синтеза всех специфических веществ организма, поддержания его высокоупорядоченной организации, активного транспорта веществ внутри клеток, из одних клеток в другие, из одной части организма в другую, для передачи нервных импульсов, передвижения организмов, поддержания постоянной температуры тела (у птиц и млекопитающих) и для других целей.
Обмен веществ (метаболизм) – совокупность биохимических реакций, протекающих в клетке и обеспечивающих процессы ее жизнедеятельности.
В ходе превращения веществ в клетках образуются конечные продукты обмена, которые могут быть токсичными для организма и выводятся из него (например, аммиак). Таким образом, все живые организмы постоянно потребляют из окружающей среды определенные вещества, преобразуют их и выделяют в среду конечные продукты.
В зависимости от общей направленности процессов выделяют катаболизм и анаболизм.
Анаболизм (ассимиляция) – совокупность химических процессов, направленных на образование и обновление структурных частей клеток, этот процесс имеет второе название – пластический обмен.
Фотосинтез: 6Н2О + 6СО2 → С6Н12О6 + 6СО2 ↑
Сюда можно отнести, например, фиксацию азота и биосинтез белка, синтез углеводов из углекислого газа и воды в ходе фотосинтеза, синтез полисахаридов, липидов, нуклеотидов, ДНК, РНК и других веществ. Анаболизм является созидательным этапом обмена веществ, он всегда осуществляется с потреблением энергии и с участием ферментов.
Катаболизм (диссимиляция) – совокупность реакций, в которых происходит распад крупных органических молекул до простых соединений с одновременным высвобождением энергии.
Катаболизм обеспечивает энергией все процессы, протекающие в клетке, и имеет второе название – энергетический обмен.
Дыхание: С6Н12О6 + 6СО2 → 6Н2О + 6СО2 + АТФ
При разрыве химических связей молекул органические соединения энергии высвобождаются и запасаются главным образом в виде молекул аденозинтрифосфорной кислоты – АТФ, универсального источника энергии у всех живых организмов (Рис. 3).

Рис. 3. Строение молекулы АТФ
По своей химической природе АТФ является мононуклеотидом и состоит из азотистого основания аденина, углевода рибозы и трех остатков фосфорной кислоты, соединенных между собой макроэргическими связями. Выделение энергии в клетке происходит при отделении одного из фосфорных остатков от молекулы АТФ, разрыв этой связи высвобождает 7,3 килокалории, тогда как при разрыве химических связей других соединений энергии выделяется в три-четыре раза меньше. При этом образуется молекула аденозиндифосфата – АДФ, с двумя фосфорными остатками. Она легко может восстановиться до АТФ, присоединив один остаток фосфорной кислоты или отдать еще один фосфорный остаток и превратиться в аденозинмонофосфат – АМФ (Рис. 4).

Рис. 4. Выработка энергии в клетке
Переход АТФ в АДФ и обратно – это основной механизм выработки энергии в клетке. Отщепление от АТФ и АДФ фосфорного остатка приводит к выделению энергии, а присоединение к АМФ и АДФ фосфорного остатка приводит к накоплению энергии.
ТЕМА: ФОТОСИНТЕЗ.
Фотосинтез – это процесс образования органических веществ в зелёных растениях. Фотосинтез создал всю массу растений на Земле и обогатил атмосферу кислородом.
КАК ПИТАЕТСЯ РАСТЕНИЕ?
Раньше люди были уверены, что все вещества для своего питания растения берут из почвы. Но один опыт показал, что это не так.
В горшок с землёй было посажено дерево. При этом измерили массу и земли, и дерева. Когда через несколько лет снова взвесили то и другое, оказалось, что масса земли уменьшилась всего на несколько граммов, а масса растения увеличилась на много килограмм.
В почву вносили только воду. Откуда же взялись эти килограммы растительной массы?
Из воздуха. Все органические вещества растений созданы из углекислого газа атмосферы и почвенной воды.
ЭНЕРГИЯ
Животные и человек питаются растениями, чтобы получить энергию для жизни. Эта энергия содержится в химических связях органических веществ. Откуда она там?
Известно, что растение не может нормально расти без света. Свет и является энергией, с помощью которой растение строит органические вещества своего тела.
Не важно какой это свет, солнечный или электрический. Любой луч света несёт энергию, которая становится энергией химический связей и как клей удерживает атомы в больших молекулах органических веществ.
ГДЕ ИДЁТ ФОТОСИНТЕЗ
Фотосинтез проходит только в зелёных частях растений, а точнее, в особых органах растительных клеток – хлоропластах.

Хлоропласты являются разновидностью пластид. Они всегда зелёные, т. к. содержат вещество зелёного цвета – хлорофилл.
Хлоропласт отделён от остального объёма клетки мембраной и имеет вид зёрнышка. Внутреннее пространство хлоропласта называется стромой. В ней и начинаются процессы фотосинтеза.

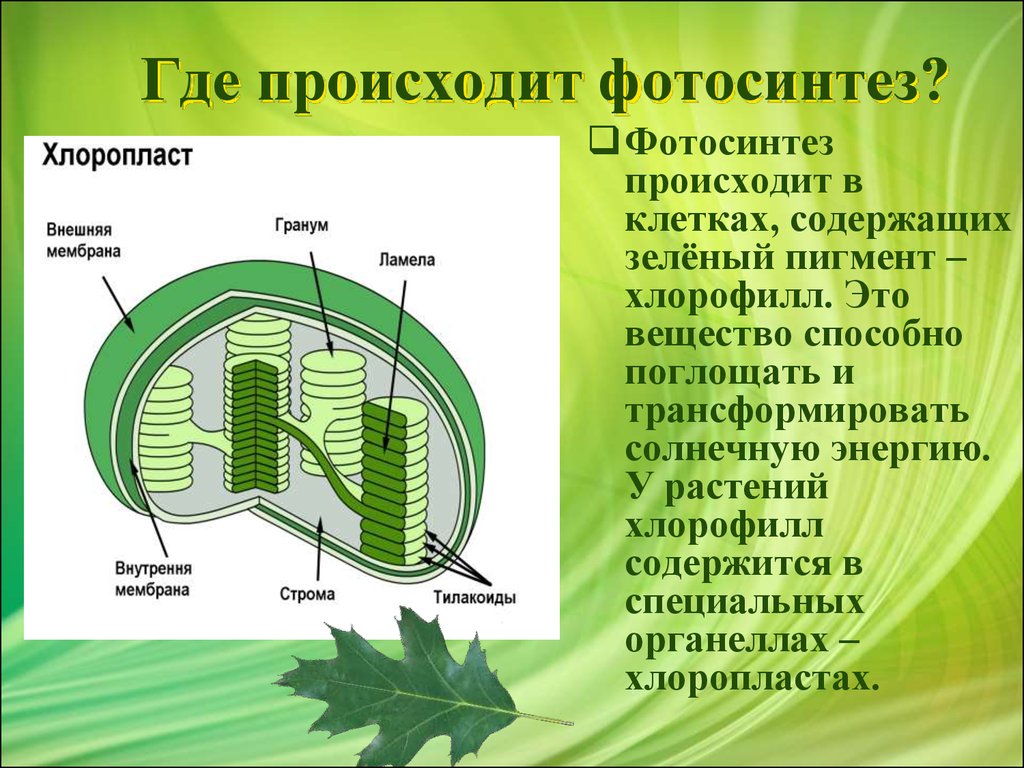
Хлоропласты являются как бы фабрикой, на которую поступает сырьё:
- углекислый газ (формула – СО₂);
- вода (Н₂О).
Вода поступает из корней, а углекислый газ – из атмосферы через особые отверстия в листьях-устьица. Свет является энергией для работы фабрики, а полученные органические вещества – продукцией.
Сначала производятся углеводы (глюкоза), но впоследствии из них образуется множество веществ разнообразных запахов и вкусов, которые так любят животные и люди.
Из хлоропластов полученные вещества транспортируются в разные органы растения, где откладываются в запас, либо используются для процессов жизнедеятельности.
РЕАКЦИЯ ФОТОСИНТЕЗА
В общем виде уравнение фотосинтеза выглядит так:
СО₂(углекислый газ) + Н₂О(вода) = органические вещества(глюкоза) + О₂ (кислород)
Зелёные растения входят в группу автотрофов (в переводе – «сам питаюсь») – организмов, которым для получения энергии не нужны другие организмы.
Основная функция фотосинтеза – создание органических веществ, из которых строится тело растений.
Выделение кислорода – побочный эффект процесса.
ЗНАЧЕНИЕ ФОТОСИНТЕЗА
Роль фотосинтеза в природе чрезвычайно велика. Благодаря ему создан весь растительный мир и озоновый экран нашей планеты.
Благодаря фотосинтезу растения:
- являются источником кислорода для атмосферы;
- переводят энергию солнца в доступную для животных и человека форму.
Жизнь на Земле стала возможной при накоплении достаточного количества кислорода в атмосфере. Ни человек, ни животные не смогли бы жить в те далёкие времена, когда его не было, или было мало.
КАКАЯ НАУКА ИЗУЧАЕТ ПРОЦЕСС ФОТОСИНТЕЗА
Фотосинтез изучают разные науки, но больше всего ботаника и физиология растений.
Ботаника – это наука о растениях и, поэтому изучает его как важный жизненный процесс растений.
 Наиболее подробно изучает фотосинтез физиология растений. Учёные-физиологи определили, что этот процесс сложный и имеет стадии:
Наиболее подробно изучает фотосинтез физиология растений. Учёные-физиологи определили, что этот процесс сложный и имеет стадии:
- световую;
- темновую.
Это значит, что фотосинтез начинается на свету, но заканчивается в темноте.
ГРУППА 306 Биология 20
ТЕМА 20: Регуляция работы генов у прокариот и эукариот.
Регуляция активности генов. Опероны у бактерий
Все клетки любого организма, какие бы функции они ни выполняли, имеют полный набор свойственных данному организму генов. Вместе с тем хорошо известно, что у любого организма клетки разных тканей и органов отличаются поразличным признакам и набору имеющихся в них белков. Даже в одной клетке на разных стадиях ее развития синтезируются и функционируют разные белки. Следовательно, располагая полной генетической информацией, каждая клетка на определенном этапе развития использует лишь ту ее часть, которая необходима в настоящий момент, транскрибируются только те гены, продукты которых нужны клетке в данный момент для осуществления ее функций. Следовательно, клетка должна располагать механизмами, определяющими, какие гены и в какой последовательности должны экспрессироваться ("выражаться", то есть давать продукт — РНК или белок). Наиболее полно регуляция генной активности изучена на примерах адаптивного синтеза ферментов у прокариот.
В зависимости от условий количество определенного фермента в бактериальной клетке может существенно изменяться. Некоторые ферменты, необходимые бактерии для усвоения определенных питательных веществ, активно синтезируются в клетке только тогда, когда эти вещества присутствуют в культурной среде, и синтез их прекращается, если каким-либо образом они удаляются из среды. Такой тип регуляции синтеза фермента называется индукцией, а вещество, включающее экспрессию гена — индуктором.
Активация и репрессия оперонов у бактерий
лактозный оперон
Один из наиболее наглядных примеров данного типа регуляции — лактозный оперон кишечной палочки — группа генов, контролирующая синтез ферментов, осуществляющих катаболизм (расщепление) молочного сахара — лактозы. Буквально через несколько минут после добавления в питательную среду для кишечной палочки лактозы бактерии начинают вырабатывать три фермента: галактозидпермеазу, бета-галактозидазу и галактозидтрансацетилазу. Как только ресурсы лактозы в среде исчерпываются, синтез ферментов сразу же прекращается.
Приведенный пример станет более понятным при рассмотрении схемы работы лактозного оперона, изучение которого позволило французским ученым Ф. Жакобу и Ж. Моно разработать собственно концепцию оперона и выяснить основные принципы регуляции транскрипции у прокариотов.
Оперон — это группа генов прокариот, находящихся под общим промотором. Все эти гены транскрибируются на одну общую молекулу мРНК. Такая мРНК, содержащая информацию о нескольких белках, называется полицистронной. Участок ДНК или РНК, содержащий информацию об одном белке, называется цистроном.
Лактозный оперон начинается с регуляторного участка, предназначенного для присоединения белка-активатора, в свою очередь необходимого для присоединения к следующему за этим участком промотору (П) РНК-полимеразы. Последовательность нуклеотидов промотора узнаётся РНК-полимеразой.
С промотором перекрывается следующий участок — оператор (О). С ним может связываться регуляторный белок-репрессор. Репрессор блокирует промотор и тем самым предотвращает транскрипцию гена.
За оператором следуют структурные гены для трех упомянутых ранее ферментов. Заканчивается оперон терминатором, прекращающим продвижение РНК-полимеразы и транскрипцию оперона.
Регуляторный белок-репрессор в незначительном количестве синтезируется в клетке постоянно, так что в цитоплазме одновременно присутствует не более 10 его молекул. Этот белок обладает сродством к последовательности нуклеотидов в области оператора и сродством к лактозе.
В отсутствие лактозы белок-репрессор связывается с операторным участком и препятствует продвижению по ДНК РНК-полимеразы: не синтезируется мРНК, не синтезируются и ферменты. После добавления в среду лактозы белок-репрессор связывается с нею быстрее, чем с операторным участком. В результате последний остается свободным и не препятствует продвижению РНК-полимеразы. Идет транскрипция и трансляция. Синтезирующиеся ферменты осуществляют транспорт в клетку и расщепление лактозы. После того как вся лактоза будет израсходована, нечем станет связывать белок-реп рессор и он снова свяжется с оператором, прекратив транскрипцию оперона. Таким образом, индукция оперона вызывается тем, что регуляторный белок не прикрепляется к оператору. Такой тип индукции называется негативным.
Параллельно наблюдается и другой тип регуляции — позитивная регуляция. При глюкозном голодании в клетке из АТФ образуется сигнальное вещество цАМФ, которое связывается с белком-активатором (САР), после чего последний приобретает способность связывать ДНК в промоторной области и усиливать транскрипцию лактозного оперона. Таким образом, когда не хватает глюкозы, стимулируется всасывание и катаболизм лактозы. При одновременном присутствии глюкозы и лактозы последняя не метаболизируется, пока существенно не упадёт концентрация глюкозы.
триптофановый оперон
В случае индукции лактозного оперона аллолактоза (индуктор) препятствует присоединению белка-репрессора к оператору, то возможен и другой вариант регуляции, когда, наоборот, индуктор придает регуляторному белку способность присоединяться к оператору. Если в первом случае соединение индуктора с белком-регулятором разрешало транскрипцию, то во втором оно запрещает ее.
Примером такой регуляции может служить хорошо изученный триптофановый оперон кишечной палочки. В его состав входят пять структурных генов, обеспечивающих синтез аминокислоты триптофана, оператор и промотор. Репрессор синтезируется вне триптофанового оперона. Пока клетка успевает расходовать весь синтезирующийся триптофан, оперон работает, синтез триптофана продолжается. Если же в клетке появляется избыток триптофана, он соединяется с репрессором и изменяет его таким образом, что этот белок связывается с оператором. Комплекс репрессора с триптофаном взаимодействует с оператором и препятствует транскрипции структурных генов, вследствие чего синтез триптофана прекращается. В отсутствие триптофана репрессор лишается способности связываться с оператором, и происходит транскрипция структурных генов оперона и, в итоге, синтез триптофана в клетке.
Другой уровень регуляции триптофанового оперона включает аттенуацию — тонкую подстройку количества продукта в зависимости от концентрации присутствующего триптофана.
регуляция генов бактериофагов
Описанные типы регуляций характеризуют механизмы регуляции отдельных оперонов, практически не касаясь регуляции экспрессии генома в целом, в то время как совершенно очевидно, что регуляция разных оперонов должна носить согласованный характер. Такой согласованный характер работы разных оперонов и генов получил у вирусов и фагов название каскадной регуляции. Согласно принципу каскадной регуляции сначала происходит транскрипция «предранних», затем «ранних» и наконец «поздних» генов в зависимости от того, какие белки требуются на разных стадиях вирусной (фаговой) инфекции.
Конечно, принцип каскадной регуляции у фагов относится к наиболее простым. У более сложно организованных организмов для осуществления большого количества функций, происходящих одновременно или с определенной последовательностью, необходима согласованная работа многих генов и оперонов. Особенно это касается эукариотов, отличающихся не только более сложной организацией генома, но и многими другими особенностями механизмов регуляции генной активности.
регуляция генов эукариот
По принципам регуляции гены эукариотов можно условно разделить на три группы: 1) функционирующие во всех клетках организма; 2) функционирующие только в тканях одного типа; 3) обеспечивающие выполнение специализированными клетками конкретных функций.
Механизмы регуляции экспрессии генов у эукариот:
- У эукариот известна регуляция генной активности на уровне структуры хроматина. В регуляции генов эукариот важную роль играют гистоны — основные белки, входящие в состав хромосом. Одни модификации гистонов характерны для активно работающих генов, другие — для молчащих. Существуют также вариантные гистоны, закодированные особыми генами. Они могут заменять "обычные" гистоны в определенных хроматиновых контекстах, влияя на регуляцию генов.
На электронных микрофотографиях в ядрах неделящихся клеток эукариот видны более плотно упакованные участки — гетерохроматин — и более рыхлые участки, называемые эухроматином. Гетерохроматин содержит центромерные и теломерные участки хромосом (облигатный гетерохроматин) и молчащие гены (факультативный гетерохроматин), а эухроматин — активно работающие гены. - Также распространенным типом регуляции экспрессии генов у эукариот является метилирование ДНК, в основном по 5 положению цитозина. Метилированная ДНК, как правило, присутствует в выключенных генах. Этим, в частности, объясняется трудность организменного клонирования, связанная с тем, что в соматических клетках (клетках тела) многие гены метилированы, поэтому когда ядром соматической клетки замещают ядро зиготы, экспрессия этих генов зачастую не активируется, т.к. метилирование генов не снимается. Надо отметить, что регуляторное метилирование распространено не у всех эукариот, например, оно не характерно для генома мушки дрозофилы.
ГРУППА 408 БИОЛОГИЯ 27,28
ТЕМА 27, 28: РЕШЕНИЕ ЗАДАЧ ПО ГЕНЕТИКЕ (ПОВТОРЕНИЕ)
Задача 1. Дано: кареглазый мужчина женился на голубоглазой женщине. У них родился голубоглазый ребёнок. Определите генотипы родителей и вероятность рождения ребёнка с карими глазами.
Решение: в задаче идёт речь об одном признаке – цвете глаз. Значит, это задача на моногибридное скрещивание. Выбираем буквенные обозначения для разного состояния генов, то есть для каждого цвета глаз. При этом учитываем, что ген кареглазости доминантный, а ген, обуславливающий голубой цвет глаз рецессивный.
Пускай А обозначает карие глаза, а ген а – голубые.
Записываем ход скрещивания условными обозначениями.
Родители Р (ставим точку и двоеточие). Обратите внимание и запомните – на первом месте всегда записывается генотип женской особи. Если поставите мужскую – это будет ошибкой. Итак, женщина голубоглазая, значит у нас может быть только один вариант генотипа – аа. То есть, женский организм – рецессивная гомозигота по данному признаку. Кареглазый мужчина может быть, как доминантной гомозиготой, так и гетерозиготой. Но так как по условию задачи у этой пары рождается голубоглазый ребёнок, значит у мужчины в гаметах будет рецессивный ген а. Записываем гетерозиготу – Аа.
Далее определяем гаметы, которые будут давать родительские формы: у матери все они будут содержать а, так как гомозиготы не дают расщепления в потомстве. Будьте здесь внимательны. Записывать нужно только тип гамет. У нас он один. А у гетерозиготного отца образуется два типа гамет: одни будут содержать А – ген кареглазости, другие – а – ген, обуславливающий голубой цвет глаз.
Определяем возможные генотипы и фенотипы детей. При слиянии сперматозоида, содержащего доминантный ген А и яйцеклетки, содержащей рецессивный ген а, развивается гетерозиготный ребёнок по данному признаку с карими глазами. Это один вариант. При слиянии половых клеток с одинаковыми рецессивными генами, будет развиваться гомозиготный ребёнок по данному признаку с голубым цветом глаз. Это второй вариант. Других генотипов у гибридов первого поколения быть не может. Значит в потомстве получаем расщепление, обусловленное гетерозиготностью отца. 50% кареглазых и 50% голубоглазых детей.
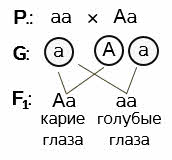
Ответ: мужчина гетерозиготен по данному признаку. Женщина - рецессивная гомозигота. Вероятность рождения в семье ребёнка с карими глазами – 50%.
Задача 2. Полидактилия у человека является доминантным признаком, а нормальное строение кистей рук – признак рецессивный. От брака мужчины, имеющего нормальное строение рук с гетерозиготной шестипалой женщиной, родились два ребёнка: пятипалый и шестипалый. Каков генотип этих детей?
Решение: обозначаем буквами доминантный и рецессивный признаки. А – полидактилия, а – нормальное строение кистей рук.
Записываем генотипы родителей. По условию задачи, женщина гетерозиготна. У мужчины известен только фенотип. Но, поскольку он имеет нормальное строение кистей рук, а ген, отвечающий за такое строение рецессивен, делаем вывод о том, что в генотипе мужчины отсутствует доминантный ген шестипалости и он является рецессивной гомозиготой по данному признаку.
Поскольку мать гетерозиготна – она даёт два типа гамет: А и а. Гомогаметный отец – один тип гамет – а.
Рассматриваем варианты слияния гамет. А и а – в результате шестипалый гетерозиготный ребёнок. Вариант а и а даст начало развитию ребёнка с нормальным строением кистей рук. Его генотип – рецессивная гомозигота. Проверяем, соблюдается ли условие задачи. Соблюдается.
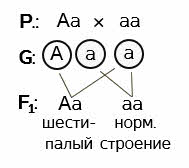
Ответ: пятипалый ребёнок – рецессивная гомозигота по данному признаку, шестипалый – гетерозиготен.
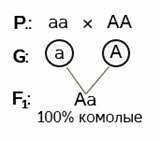
Задача 3. От скрещивания комолого быка айширской породы с рогатыми коровами в первом поколении получили 18 комолых (безрогих) телят. Во втором поколении гибридов родилось 96 телят. Часть из них безрогие, а часть рогатые. Определите количество комолых телят во втором поколении и запишите ход скрещивания.
Решение: ген комолости доминирует над геном рогатости. Даже если нам это неизвестно – такой вывод можно сделать из условия задачи. В первом поколении наблюдается единообразие гибридов. А во втором происходит расщепление с проявлением признака, кодируемого рецессивным геном (второй закон Менделя). Делаем вывод о том, что ген комолости – доминантный, а ген, определяющий наличие рогов – рецессивный.
Записываем условие. Рогатые коровы могут иметь только один генотип – рецессивные гомозиготы. Комолые быки могут быть как гомозиготными, так и гетерозиготными. Но поскольку в первом поколении гибридов рецессивный признак не проявляется (18 телят – достаточное количество, чтобы это предположить с большой долей вероятности), значит мужская особь – доминантная гомозигота.
Гомозиготы дают по одному типу гамет.
В результате оплодотворения образуется единообразное потомство. 100% гетерозигот.
Скрещиваем между собой гибридов первого поколения. Родительские особи дают по два типа гамет. Для того, чтобы рассмотреть все варианты их слияния – построим решётку Пеннета. Помните? Женские гаметы записываются вертикально, а мужские горизонтально. Заполняем решётку.
Получаем расщепление по фенотипу в соотношении три к одному. Три части комолых и одна часть рогатых телят. Так как по условию, общее количество телят 96, то три части от этого количества составит 72 телёнка.
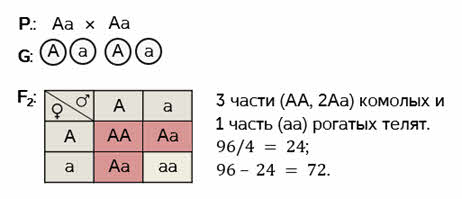
Если бы в условии задачи не было задания записать ход скрещивания, её можно было бы решить, опираясь только на второй закон Менделя. Согласно его формулировке во втором поколении гибридов наблюдается расщепление: три части особей с доминантным признаком и одна часть с рецессивным. Всё те же 72 телёнка.
Ответ: 72 телёнка.
Неполное доминирование.
Задача 1. У земляники красная окраска ягод неполно доминирует над белой. Какое потомство следует ожидать от скрещивания двух растений с розовыми ягодами? Запишите генотипы и фенотипы гибридов.
Решение: обозначим доминантный ген буквой А, а рецессивный – а. Отметим, что доминантные гомозиготы будут иметь красные ягоды, рецессивные гомозиготы – белые, а гетерозиготы – розовые.
Записываем ход скрещивания. Обе родительские формы – гетерозиготы. Так как по условию они имеют розовые ягоды. Гетерозиготы дают два типа гамет.
Строим решётку Пеннета. Определяем генотипы и фенотипы первого поколения гибридов. Обратите внимание, что при неполном доминировании (промежуточном характере наследования) расщепление по фенотипу совпадает с расщеплением по генотипу.
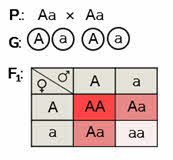
Ответ: 25 % доминантных гомозигот с красными ягодами, 50 % гетерозигот с промежуточной окраской плодов – розовой и 25 % рецессивных гомозигот с белыми ягодами.


Комментариев нет:
Отправить комментарий